|
|
Главная :: Архив статей :: :: Ссылки |
|
|
Главная :: Архив статей :: :: Ссылки |
Наши друзья Помощь сайту R935344738975 Наша кнопка Партнеры • Sunward |
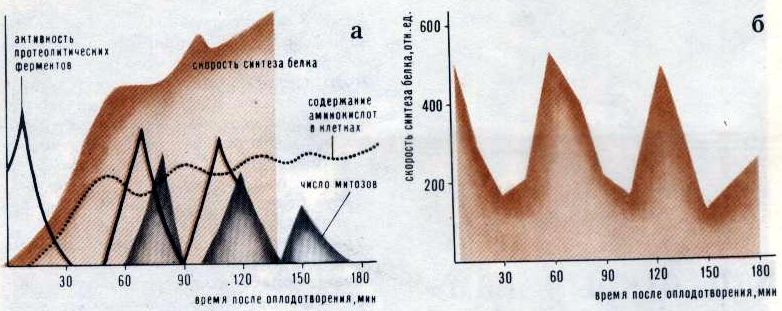
Архив статей > Биология > Кто открыл нам времени счет? Кто открыл нам времени счет? Л. Л. Литинская Живая материя, зародившись и продолжая существовать в ритмически изменяющемся мире, естественным образом унаследовала от него способность к периодическим изменениям. Любой биологический объект характеризуется целой иерархией ритмов: годовых, сезонных и суточных (или, как их принято называть, циркадных1), а также более коротких - часовых, минутных и секундных... Лучше всего исследованы циркадные, приливно-отливные и сезонные ритмы. Менее повезло ритмам с более короткими периодами. И пожалуй, позднее всего начали изучаться часовые ритмы живого хронометра: первые сведения об околочасовых изменениях активности организмов появились в 1961 году, когда было показано, что колебания сухого веса ганглиозных клеток сетчатки мыши происходят с периодом около 40 минут. С тех пор подобные ритмы были обнаружены у самых разных биологических объектов; именно об этих ритмах мы и поведем рассказ. Ритмы с периодом, близким к часу, наиболее тщательно изучил японский исследователь И. Мано на эмбрионах морского ежа. Первые публикации Мано относятся к 1968 году, хотя начались эти работы гораздо раньше. Причина молчания проста: на первых порах существование околочасовых ритмов приводило ученых в недоумение (ведь деление суток на 24 часа представляется чисто случайным приемом, не связанным с какими-либо реально наблюдаемыми природными ритмами вроде смены дня и ночи); следствием этого явилось недоверие к результатам экспериментов. В институте, где работал Мано, была создана специальная комиссия, и только после того, как эта комиссия тщательно проверила работу ученого и подтвердила его результаты, "табу" было снято и статьи Мано начали появляться в печати. Мано показал, что после оплодотворения яиц морского ежа в них возникают колебания скорости синтеза белка, активности протеолитических ферментов, проницаемости плазматической мембраны, числа полирибосом, числа SH-групп в кислоторастворимом белке и т. д., причем период этих колебаний совпадает с циклами делений клеток и составляет примерно 40 минут. Во всем этом вроде бы нет ничего противоестественного. Однако и после добавления ингибиторов ядерного и клеточного деления и после гомогенизации эмбрионов, и даже в растворе, полученном после удаления из него разрушенных клеток,- везде сохранялись все те же 40-минутные колебания (рис. 1); по данным Мано, синтез РНК и ДНК также изменяется ритмически благодаря соответствующим изменениям активности ДНК- и РНК-полимераз. То есть получается, что все компоненты эмбриона даже в изолированном состоянии следуют одному и тому же ритму. Не правда ли, тут есть чему удивиться? Казалось бы, эмбриональная клетка - особая клетка. Однако напомним, что впервые околочасовые ритмы были обнаружены в высокоспециализированных ганглиозных клетках сетчатки. Теперь же стало известно, что скорость синтеза белка, проницаемость клеточных мембран, количество РНК и белка и другие характеристики изменяются с околочасовой периодичностью во многих специализированных клетках: нервных клетках сетчатки, клетках мозжечка, секреторных клетках слюнных желез, клетках печени и поджелудочной железы. Все эти исследования были выполнены в Институте биологии развития АН СССР.
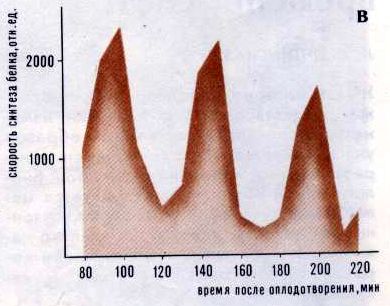
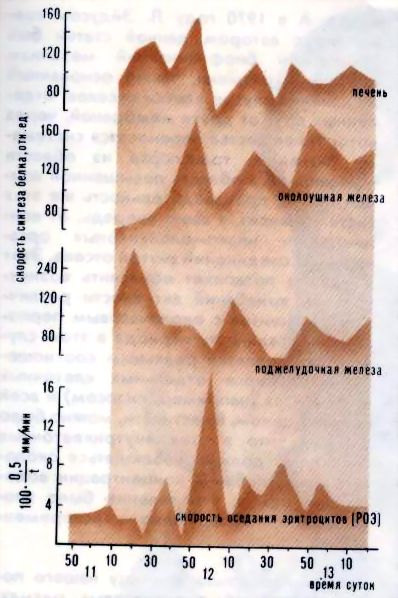
Рис. 1. Ритмические процессы, происходящие в эмбрионах морского ежа (а), гомогенате эмбрионов (б) и в надосадочной жидкости (в) Но происходят ли околочасовые колебания в клетках постоянно или же это лишь особая форма реакции клетки на внешнее физическое или химическое воздействие? Многими исследователями было установлено, что для возникновения околочасовых ритмов такое воздействие необходимо: например, в опытах Мано - это сам факт оплодотворения яиц морского ежа. Однако оставалось неясным - инициировал ли внешний агент колебания или же он лишь синхронизировал колебания, уже происходящие в различных группах клеток. Сделать выбор между этими двумя возможностями очень долго не удавалось: ситуация была похожа на классическую ситуацию в квантовой физике, когда любое измерение оказывает воздействие на объект, что приводит к изменению самих измеряемых характеристик. И все же эта деликатная задача была успешно решена: опытами по изучению ритма синтеза белка в клетках сетчатки мышей удалось доказать, что внешний агент вызывает только синхронизацию колебаний, происходивших в клетках сетчатки и прежде. Ну а связаны ли между собой околочасовые ритмы разных клеток живого организма? Синхронны ли колебания, например, в клетках печени и поджелудочной железы, в эритроцитах и секреторных клетках слюнных желез? Эксперимент показал, что ритм синтеза белка в печени, поджелудочной и слюнной железах крысы обнаруживает значительное сходство (рис. 2), хотя синтез и выделение пищеварительных ферментов в слюнной и поджелудочной железах регулируются разными способами. Злокачественные клетки ведут себя, в общем, так же, как и нормальные. Например, в клетках асцитной карциномы Эрлиха (АКЭ), развивающихся в брюшной полости мыши и в гема-цитобластах человека при остром лейкозе, были обнаружены все те же околочасовые колебания размеров и сухого веса клеток и ядер, прочности и проницаемости плазматической мембраны и т. д. Надо сказать, что когда при изучении клеток АКЭ были обнаружены околочасовые ритмы (эта работа была сделана в Институте биологической физики АН СССР в 1964 году, когда существование околочасовых ритмов у всех нормальных клеток еще не было доказано), возникла гипотеза: в раковых клетках "биологические часы" идут в 20 раз быстрее, чем в нормальных, для которых характерен циркадный, околосуточный ритм, а ускорение хода часов ведет к преждевременной гибели организма. Красиво, не правда ли? Красиво, но, увы, неверно...
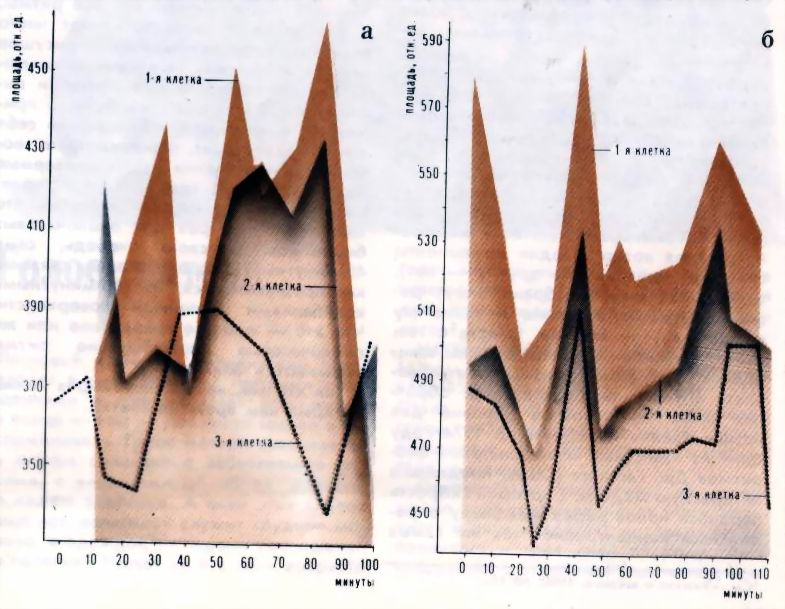
Рис. 2. Ритмы наблюдаемые в различных органах одного животного В последние годы широко используется метод культивирования клеток вне организма, на питательных средах. Оказалось, что и в клетках, растущих in vitro, сохраняются все те же околочасовые ритмы. В этом случае удалось тщательно проследить за их синхронизацией: оказалось, что в первые двое суток после посева клетки колеблются асинхронно (рис. 3), на третьи сутки между колебаниями клеток возникает некоторая связь, а на четвертые сутки синхронно колеблются 60-80% клеток (синхронизация происходила так медленно, видимо, потому, что опыт проводился при пониженной температуре). Синхронность околочасовых колебаний наблюдается даже в том случае, если клетки оказываются практически почти полностью обособленными (например, при сильном разведении клеточной суспензии). Кроме того, в каждой отдельной клетке синхронно (но не обязательно синфазно) изменяется большое число самых разных параметров. Эти колебания сохраняются и в безъядерных клетках, и даже в бесклеточных системах.
Рис. 3. Синхронизация клеток по мере роста культуры: а) - через 6 часов после пересева б) - на четвертые сутки после пересева Существенная особенность околочасовых колебаний - их устойчивость к внешним воздействиям. Так, период этих ритмов меняется лишь незначительно при изменении температуры от 18 до 37°С; правда, при 0°С колебания всегда прекращаются. На уже идущие колебания не влияет ни облучение, ни обработка клеток антибиотиками, прекращающими деление. Правда, удалось заметить изменение ритма синтеза белка в слюнных железах при увеличении или уменьшении интервалов между кормлениями животного; но это - эффект обучения, который проявляется на уровне организма. На чисто же клеточном уровне таких изменений пока получить не удалось. Но в чем же все-таки заключается причина возникновения околочасовых ритмов? Циркадные, околосуточные ритмы изучаются вот уже 250 лет. Однако причина их возникновения до сих пор так еще и не расшифрована; это служит для исследователей околочасовых колебаний немалым утешением - ведь у нас в запасе есть еще 230 лет... И все же попробуем рассказать о некоторых гипотезах и оценить их применимость к околочасовым колебаниям. Одной из первых гипотез была предложенная Ч. Эретом (США) генетическая модель биологических часов, получившая название "хронон". Отсчет времени в этой модели отождествлялся с последовательностью различных биохимических реакций на расплетающихся в ходе клеточного цикла нитях ДНК. Однако в нашем случае модель Эрета не годится, так как околочасовые ритмы сохраняются и в системах, не содержащих ДНК. Другая гипотеза, наиболее четко сформулированная в 1973 г. Е. Сельковым (СССР), заключается в том, что колебания возникают в однородной, гомогенной биохимической системе так же, как и в автоколебательных химических системах , в которых химические процессы протекают циклически и поэтому могут служить отметчиками времени. Биохимические колебания такого типа (с периодами около минуты) были обнаружены экспериментально в одной из ферментативных систем дрожжей; считается, что подобные колебательные процессы могут происходить и с более длительными (даже суточными) периодами. Однако неоднородность, гетерогенность структуры реальной клетки создает, на наш взгляд, условия и для протекания периодических процессов несколько иного типа. А именно: если какая-либо биохимическая реакция протекает внутри клеточного отсека, ограниченного от внешней среды полупроницаемой мембраной (то есть мембраной, через которую одни компоненты способны проникать, а другие - нет), то свойства этой мембраны могут решающим образом влиять на кинетику реакций внутри объема. Вместе с тем свойства самой мембраны должны зависеть от процессов, которыми она управляет; в результате образуется обратная связь - основное условие для возникновения колебаний. В 1974 году один из известных исследователей биоритмов Дж. Гастингс (США) предложил интересную модель, в которой скорость переноса ионов через мембрану определялась концентрацией тех же самых ионов. А в 1970 году Л. Эйдусом совместно с автором данной статьи был предложен биофизический механизм клеточной ритмики, также основанный на наличии внутри клетки отсеков, отделенных друг от друга мембраной, через которую вещества переносятся системами активного транспорта из области пониженной в область повышенной концентрации; производительность же этих систем зависит, в свою очередь, от концентрации низкомолекулярных органических соединений внутри отсека. Этот механизм позволяет объяснить возникновение колебаний активности различных ферментов с околочасовым периодом. Длительность периода в этом случае определяется реальным соотношением размеров отдельных клеточных фрагментов (например, лизосом) и всей клетки в целом. В частности, можно было ожидать, что в этих внутриклеточных структурах должны наблюдаться околочасовые колебания концентрации водородных ионов; это явление было впоследствии обнаружено экспериментально. Вот мы и подошли к концу нашего повествования об околочасовых ритмах. Существование их теперь уже не подвергается сомнению. Но причины возникновения этих ритмов все же еще не разгаданы до конца. Большинство исследователей биологических часов считают, что все ритмы, в том числе и часовые, имеют чисто внутреннее происхождение, присущее самому организму. Но нельзя исключить и возможность того, что они вызываются какими-то внешними причинами. В частности, обращает на себя внимание тот факт, что некоторые геофизические явления (так называемые микропульсации геомагнитного поля) имеют период, равный 40 минутам, что характерно как раз для околочасовых биоритмов. В свою очередь, сами 40-минутные микропульсации связаны, как предполагается, со 160-минутными колебаниями солнечной поверхности. Что это - простое совпадение или же околочасовые биологические ритмы вызываются Солнцем? Так кто же, как сказал Генрих Гейне, "открыл нам времени счет"? 1Термин "циркадный" (то есть околосуточный) не совсем правилен, так как этот ритм имеет околосуточный характер только при постоянных условиях внешней среды: при возможности свободного выбора (одна половина клетки находится на свету, другая - в темноте) животные следуют строго суточному ритму (см. "Химию и жизнь", 1976, № 1, с. 15). Похожие статьи: 1. Всё циклично в этом мире Старикович С. Химия и Жизнь №5, 1971 г., с. 40-43 2. Цепь суточных ритмов Дердиященко А. А. Химия и Жизнь №5, 1987 г., с. 44-47
Главная :: Архив статей :: :: Ссылки |